水草オタクのレンコンがたり.
寒くなってきました。
今回は、正月に欠かせない冬の作物、レンコンについて、水草オタクがみっちり語っていこうと思います。
- I. 世界のハス
- II. ハスの進化史と分類的位置
- III. ハスの草体について
- IV. レンコンの穴とハスの通気システム
- V. 葉による加圧と気体循環の制御
- VI. まだわからない部分もある、レンコンの構造
I. 世界のハス
レンコンはハスの肥大した地下茎です。では、ハスの仲間は世界に何種あるかと言えば、2種しかいないと2024年現在では考えられています。アジアからオーストラリアに分布するハスNelumbo nuciferaと、北米(~南米北部?)に分布するキバナハスNelumbo luteaです。キバナハスは花が黄色であるだけでなく、草体は概して通常のハスよりは小柄で葉柄に棘がなく、花弁数が少なく葉色が濃い点で区別できます。栽培に関して言えば、キバナハスの方が難しい傾向があるようです。ハスとキバナハスは古生物学的にはかなり昔に分岐したと考えられてきましたが、分子生物学的には更新世ごろ、もしくはそれ以降に分岐したもののようです。
ハスはインドおよび中国において古くから利用され、東洋のイメージが強い植物といえます。ハスとスイレンを区別する際、しばしばハスには”Indian Sacred lotus"であるとか”Indian bean"、”Chinese water lily"と呼ばれるのはその傍証でしょう。ヨーロッパにハス類の分布はなく、Nelumboという属名はシンハラ語に由来するといわれています。
アジアにおけるハスの利用および改良は、どうも中国のものと南~東南アジアのものに分けて考えた方がよいように思われます。アジアのハスは温帯性ハスと熱帯性ハスの2系統に分けられ、今回紹介するレンコンがとれるのは温帯性ハスのみです。
熱帯性ハスは細いランナー状の根茎が主体で冬場の根茎の肥大は顕著でなく、同じ条件で育てた場合には熱帯性ハスのほうが2~3か月長い開花期間を持ち、常緑性の傾向があります。国内で見かけることは殆どありませんが、タイやベトナムでは肥大しない根茎を食用にします。日本においてもベトナム料理店には稀ながらも熱帯性ハスの根茎が売られています。熱帯のハスはレンコンを作らないことや、またインド(特に南部)では神聖視されて食べない、仏教の影響が強い地域でも神聖視される面が強い・・・などから、食用としての改良は南・東南アジアでは盛んではないようです。そのため研究が遅れており、どのあたりの地域まで熱帯性ハスがいるのか、熱帯性ハスの中での遺伝的分化はどの程度進んでいるのか、興味深いところです。
温帯性ハスは根茎用ハス、種子用ハス、花ハスの3つに分けられることが多いですが、それらはしばしば多系統であるようです。花ハスはおもに中国で発展した栽培品で、草体は小型ながらも花が高くあがります。種子利用ハスの分類ははっきりしませんが、(多系統ともされる)一般に栽培の中心は根茎利用ハスより南方に位置します(湖南省、江西省、福建省、ベトナム)。これらの種子利用ハスは”亜熱帯性の”分布を示すように思われますが、今後の研究に期待したいところですし、もし可能であれば各品種を実際に栽培して形質を比較したいところです。
今回の主役である根茎利用ハスもまた(日本で利用されるものを含めて)中国由来の品種であり、一般的には白花で開花数は少なく、草体は大型である傾向があります。根茎利用ハスは日本においても中国においても最も作付け面積の広いハスであり、レンコンは日本、中国ともに一般的な食材です。中国においては湖北省、江蘇省、安徽省、浙江省で栽培されており、日本では茨城県が生産量の半数近くを占めています。
日本で育てられている花ハスおよびレンコン用の食用蓮、繊維用ハスはすべて温帯性ハスおよび、温帯性ハスとキバナハスの交雑に由来します(すくなくとも温帯性ハスとキバナハスは交配可能で交配種にも稔性があります)。さらに、食用ハス、花ハスともに主要な品種は中国に由来し、日本由来のハスはごく一部の”古代ハス”と呼ばれるような個体群などを除けばほぼ栽培されていません。
西洋およびオリエントにおけるハスの存在感はほとんどないのですが、エジプトではかつて珍重された経緯があります。スイレン類(Nymphaea nouchali var. caeruleaとNymphaea lotus)が古典的に利用されていたエジプトにハスを持ち込んだのはおそらくアケメネス朝ペルシア帝国で、前5世紀ごろから用いられるようになったようです。エジプトのハスは自然分布ではありませんが、エジプトにハスが持ち込まれるとイシス教において神聖視され、古来より用いられてきたスイレン類にとってかわったようです。但し、エジプトに移植されたハスは現存しておらず、スイレンと似る点も多いことから記述および当時の絵に頼るほかありません。
II. ハスの進化史と分類的位置
ハスは生きた化石と言えます。双子葉植物のなかでも特に原始的な植物であり、ハス科植物はハス属以外の属は現存していません。ハス科は真正双子葉植物の中でも最も早期に分岐したヤマモガシ目に属しますが、ほかのヤマモガシ目植物とは白亜紀前期の時点ですでに分かれていたと考えられています。最近記載されたNotocyamus hydrophobusは1億2100万年前の白亜紀アプチアンから産出しており、最初のハス科植物ながら白亜紀のハス科植物としては全草に近い保存をされており、現在のハスのものに似た花托がみられ、すでに水生であったと考えられています。ハス属とされる化石は約1億1000万年前のアルビアンから産出するN. lusitanicaと N. weymouthiが最初で、双子葉植物の属としては最も古い部類に入ります。白亜紀からは知る限りで5属22種のハス科植物が知られており、ハス属としてはN. choffati, N. kempii, N. intermedia, N. primaeva, N. laraimiensis, N. tenuifolia, N. lakesiana, N. crossii, N. dawsoni, N. puertae, N. provinciale, N. arctica, N. amurensis, N. orientalis, N. schweinfurthi、ハス属以外としてはNelumbites virginiensis、Paleonelumbo macroloba, Nelumbago peltata, Exnelumbites callejasiae, Notocyamus hydrophobusが挙げられます。これらの多くは葉のみから記載されていますが、現在のハスと比較すると葉中央の円盤状部分を欠いていたり(このことはのちに述べるが生理的に極めて重要な点と言える)、葉に鋸歯を持っていたりするなど、現在のハスと祖先を同じくするものの、現在のハスの祖先というわけではないものが多いようです。ハス科植物は白亜紀の時点では少なくとも北米、ユーラシア、アフリカ、南米に分布し、ほぼ世界中と言っていい分布を確立したと考えられます。ハス科植物の繁栄は隕石衝突による白亜紀末の大量絶滅後も(少なくとも北半球では)続き、N. protolutea, N. aureavallis, N. changchangensis, N. nagalensis, N. nipponica, N. weylandi, N. protospeciosa, N. endoana, N. hungarica, N. lacunosa, N. megalopolitana, N. minimaなどたくさんの種が記載されています。しかし第四期の氷河期により分布は著しく縮小し、アジアと北米にしか生き残れなかったと考えられています。ハスは寒さにそこまで強い植物ではなく、レンコンの保存適温も5度以上、5~8度が適温とされています。
III. ハスの草体について
ハスの草体は横走する地下茎(肥大化したものが所謂レンコン)と、その節から出る根、葉、花茎から構成されます。葉は各節から1枚ずつでるように見えますが、基部には発達しない、葉鞘状の低出葉Cataphyllが2枚あるため、各節から出る葉は3枚ずつであると考えられています。葉は盾状(Peltate)と表現される、長い葉柄が円形ないしやや漏斗状になった葉の中央下部につく形態をしており、葉柄には棘があります。(キバナハスにはない。)葉柄の断面には4つの大きな穴および、5つの小さな穴があり、これらはGas Canalと呼ばれます。ハスは地下茎の節から発根し、”レンコン”の側面にあたる節間からは殆ど発根しません。地下茎の節には3つの小室があり、節間にあたる”レンコン”にもGas canal・・・すなわち穴が約10個あります。花は1つずつつく両性花で、放射相称性を示します。花は直径10~30㎝ほどのものが多く、ハスの場合白色~桃色です。花は水面より高くつくものの、葉柄と花柄はほぼ同長か花柄の方が短いことが多く、葉より下に花がみられることもしばしばです。花の萼片や花弁の数は可変が激しく、萼片数は2~5、花弁数は20~30、雄蕊数は200~300とされます。雌蕊は20~30心皮で大型化した花托に埋もれます。この花は発熱することが知られており、また雌蕊の構造も興味深いのですが今回はレンコンの話なのでこの辺りにしておきましょう。
IV. レンコンの穴とハスの通気システム
ここからが本題です。
レンコンの穴はしばしば「通気のため」とされ、また葉柄の穴に関しても水を通すハスの葉シャワーや夏休みの自由研究などの題材にしばしばされてきました。しかしながらレンコンの穴とそれを介したハスの通気システムは植物の中でも極めて特殊かつ複雑な構造であり、他の水生植物とは一線を画する面があります。この構造全体の把握はとても難しく、100年ほどにわたって研究されている状況です。ここからは、レンコンの穴を通じて、1億年かけて完成されたハスの”異常な”水生適応について書いていきます。
レンコンの穴は一見放射相称に見えますが、じつは上下軸が存在する左右対称な構造です。しかも、そのそれぞれの穴には特有の機能と複雑な接続関係があり、同じ穴が並んでいるだけではないのです。
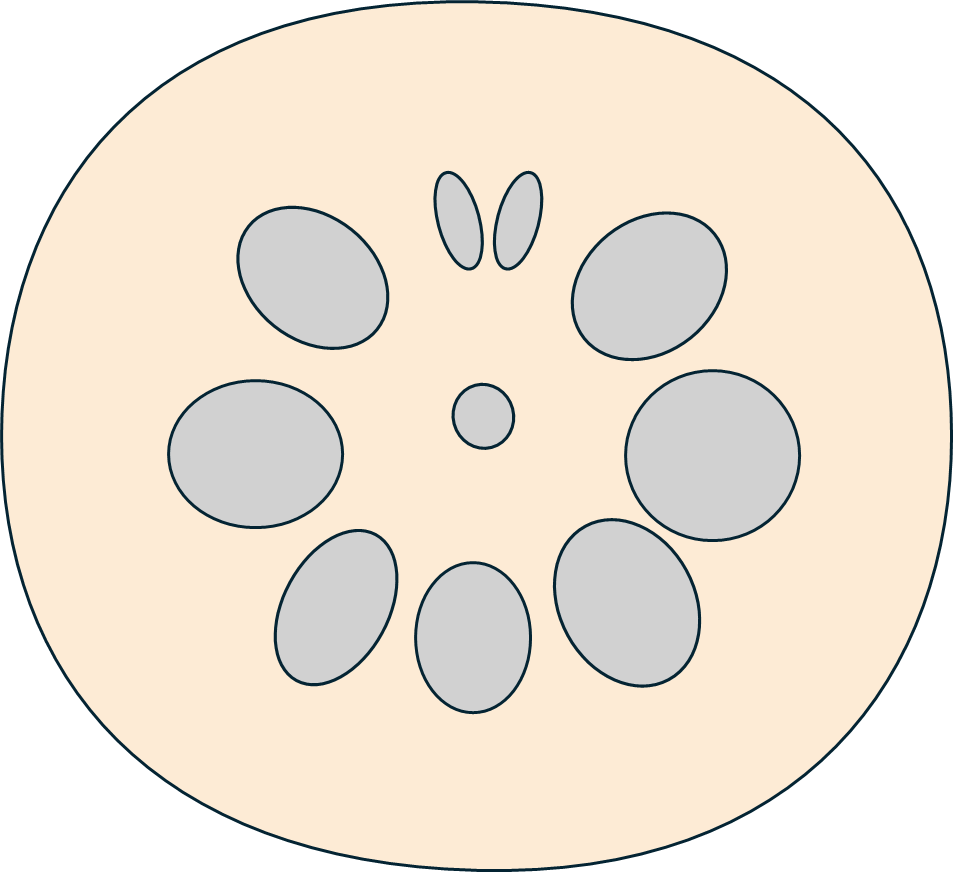
レンコンの穴がだいたい10個であることはよく知られていますが、それに上下軸があることはほとんど触れられていません。レンコンの大きな穴はだいたい10個ですが、その中にはほぼ必ず、小さい1ペアとその対極に位置する大きな1穴があります。それが軸になり、小さい1ペアが上側に位置します。
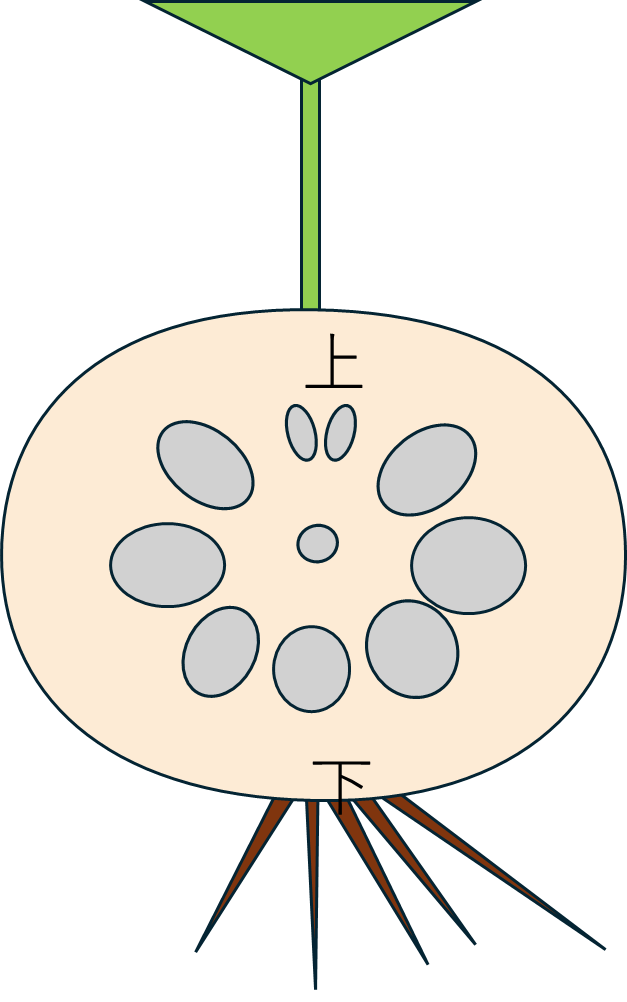
断面側から草体をみると、こんな感じになります。
次に、この穴に名前を付けます。

次に、ハスの茎の穴にも名前を付けます。ハスの茎の穴もやはり放射相称ではなく明確な極性があります。

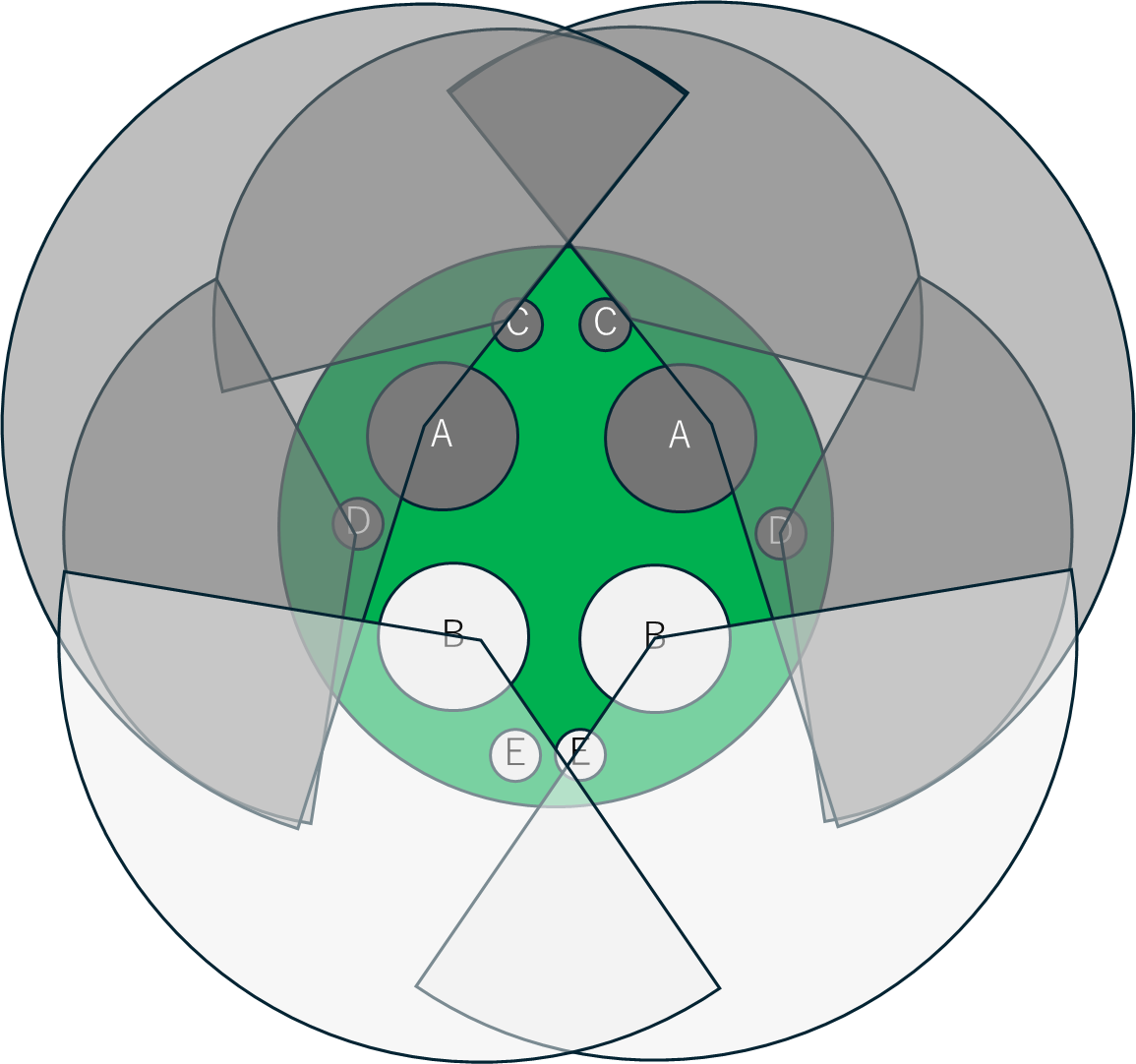
このなかで、BおよびEで示した部分からは水中で切っても気泡が出ず、A, C, Dから気泡が出ることが知られています。つまり、A, C, Dは根茎からの上向きの流れ、B、Eは葉から根茎への下向きの流れを担当しています。ハスの通気システムは基本的に各穴を流れる気体の流れは一方向性かつ規則的で、ほかの水生植物に見られるものとは明らかに異なっています。
これらの穴が節においてどのように接続しているかについて模式的に図示します。レンコンの節はなかなか観察しにくく、シリコン型をとって研究が行われています。節には3つの小室があり、X, Y, Zで示しています。

Matthews, P. G., & Seymour, R. S. (2006)より作図。
葉から送られてきた空気は小室Zを通ってレンコンの側方2対にあたるIIおよびIIIを通って草体の前後に配分されます。IIを通る空気の一部は小室Xに接続して上側に位置するIに入り、Iはさらに下流側で小室Yを介して葉柄のA, C, Dに接続し葉に送られます。
きわめてややこしいので、1方向にするとこのような図になります。

ハスの葉柄はガスの移動する極性が1本の葉柄の中でも分業していることが知られていますが、こうしてみると基本的には新しい葉から地下茎を2節通って古い葉に流しているように見えます。水中に空気を効率的に送るには、入り口と出口がある一方向性の経路が必要です。特に大型化する水生植物、とくに深い水から抽水・浮葉するためにはその空気輸送距離はそうとうに長いものになり、効率的な輸送システムが必要となります。ただ穴をあけているだけでは不十分なのです。他の浮葉性水草(たとえばコウホネやスイレン、アサザなど)では、新しい葉から入ったガスを地下茎に通し、古い葉から排出するシステムが用いられていますが、基本的にはこれに似た、しかしより複雑なシステムといえます。
しかし、それで終わらないのがハスです。
レンコンの謎が少し解けましたが、その先を見るにはハスの葉に目を向けねばなりません。
V. 葉による加圧と気体循環の制御
ハスの葉の中心に溜まった水から、ブクブクと気泡が出てくることはかなり前から知られてきましたし、気にしてみるとしばしば見かける光景でもあります。
ハスの葉の中央部には白っぽい円盤状の部分があり、これが地下茎から葉柄を伝って出てきた空気の出口となっています。この部分(Central plate)には特殊な大型の気孔があり、根茎からでたガスの出口となっています。ハスのCentral plateの気孔は午前中に開き、正午に閉じ、午後に再び開くという周期的な動きをしており、草体内のガス需要や、また草体から流出するガス流量、ないし草体内で生じたCO2の光合成への利用が制御されていると見られます。
Central discに接続するのは3系統ある葉柄を上行するGas canalのうち最も太いAの一部のみで、残りは一旦集まったのちに葉脈に沿って広がります。逆に、葉脈に沿った通道組織はBとEにもまとめられ、葉柄を下行します。傘状の葉にどのGas canalがつながるかは葉の部位によっても異なります。概して、葉の2/3が排出につながるGas canalに、1/3が空気の取り込みにつながるGas canalに向かうようです。

Vogel, S. (2004).より作図。
Vogel, S. (2004).を参考に作図。
さて、ここまでハスの発達した”一方向性の”ガス循環についてみてきましたが、その駆動力についても触れてみます。
ハスの植物体内を通るガスは光合成ガスではなく、大気中の組成とほぼ変わらないガスが送られていることがわかっています。
ハスの植物体内へのガス取り込みは気孔を介しており、ハスの気孔はまるで浮葉植物のように上面のみに分布し下部や葉柄、花柄にはほぼ欠落しています。(スイレンと違って抽水植物なのでハスの葉には下部にも気孔がある、とする誤解が散見されるので注意。)気孔は完全に閉じることができず、常にある程度開口しています。
気孔から入った空気がハスの葉で加圧され、日光による熱によりその加圧が推進される(Thermo-osmosis)ことはよく知られていますが、これはあくまでも加熱により加圧が起きるのであって、光合成による影響ではないことがわかっています。ハスの葉の索状組織の下には密な柔組織があり、その下に空気の入った小室がみられます。小室は葉の断面の半分以上を占め、小室内で蒸発した水分による葉内外の水蒸気差によって外気の乾燥した空気を取り込む(hygrometric pressurization)と考えられています。他にも、温度差があり半透膜を隔てたガスが半透膜を通って移動するKnudsen効果もその駆動力として考えられることもあります。加圧された空気は柔組織により外に逃げることが防がれ、小室につながるGas canalへと導かれてガス循環の駆動力となるようです。
VI. まだわからない部分もある、レンコンの構造
今回の記事を書くにあたってハスの構造を復習していたところ、ハスの葉柄をフィルターとして使えるという自由研究が出てきて驚かされました。
ハスの葉柄の穴にはフィルター状ないし網目状の水を弾く膜があり、この構造の意義に関してはなかなか解釈の一致を見ないところです。一つの説としては破損した茎から水が侵入するのを防ぐというものがあり、たしかに実験上は機能するようではあるそうです。同様のネット状ないしスポンジ状の構造は根茎の節部にも存在し、計算上は同様に水が地下茎に侵入することを防げるとのことですが、他にも肺のように酸素を取り込む機能など、ほかの意義がある可能性もあります。
食卓のレンコンにはまだまだ謎が沢山ありそうです。
Prigioniero, A., Scarano, P., Ruggieri, V., Marziano, M., Tartaglia, M., Sciarrillo, R., & Guarino, C. (2020). Plants named “Lotus” in antiquity: Historiography, biogeography, and ethnobotany. Harvard Papers in Botany, 25(1), 59-71.
Mekbib, Y., Huang, S. X., Ngarega, B. K., Li, Z. Z., Shi, T., Ou, K. F., ... & Yang, X. Y. (2020). The level of genetic diversity and differentiation of tropical lotus, Nelumbo nucifera Gaertn.(Nelumbonaceae) from Australia, India, and Thailand. Botanical studies, 61, 1-11.
Tian, H. L., Xue, J. H., Wen, J., Mitchell, G., & Zhou, S. L. (2008). Genetic diversity and relationships of lotus (Nelumbo) cultivars based on allozyme and ISSR markers. Scientia Horticulturae, 116(4), 421-429.
Zhou, P., Jin, Q., Qian, P., Wang, Y., Wang, X., Jiang, H., ... & Xu, Y. (2022). Genetic resources of lotus (Nelumbo) and their improvement. Ornamental Plant Research, 2(1), 1-16.
霞正一. (2003). 食用ハスの交配育種法: 遺伝資源の収集から品種登録まで.
篠原啓子, 圓藤勝義, 澤田英司, 永峰賢一, 水口博之, & 福井裕行. (2016). 食用ハス (Nelumbo nucifera) における肥大根茎 (レンコン) の形状の品種・系統間差.
Gobo, W. V., Kunzmann, L., Iannuzzi, R., Dos Santos, T. B., da Conceição, D. M., Rodrigues do Nascimento Jr, D., ... & Coiffard, C. (2023). A new remarkable Early Cretaceous nelumbonaceous fossil bridges the gap between herbaceous aquatic and woody protealeans. Scientific Reports, 13(1), 8978.
Li, Y., Smith, T., Svetlana, P., Yang, J., Jin, J. H., & Li, C. S. (2014). Paleobiogeography of the lotus plant (Nelumbonaceae: Nelumbo) and its bearing on the paleoclimatic changes. Palaeogeography, Palaeoclimatology, Palaeoecology, 399, 284-293.
Matthews, P. G., & Seymour, R. S. (2006). Anatomy of the gas canal system of Nelumbo nucifera. Aquatic Botany, 85(2), 147-154.
Vogel, S. (2004). Contributions to the functional anatomy and biology of Nelumbo nucifera (Nelumbonaceae) I. Pathways of air circulation. Plant Systematics and Evolution, 249, 9-25.
巽二郎, 野田, 千代, 山内, 章,飯嶋, 盛雄, & 矢野勝也. (1999). ハス (Nelumbo nucifera) 葉身における表面温度の分布特性. 日本作物学会東海支部会報, 15-18.
Blaylock, A. J., & Seymour, R. S. (2000). Diaphragmatic nets prevent water invasion of gas canals in Nelumbo nucifera. Aquatic Botany, 67(1), 53-59.
https://www.tsukuba.ac.jp/community/students-kagakunome/shyo-list/pdf/2018/j1.pdf