オオカナダモ
最強の水草とは何でしょうか?
物騒な言い方をしましたが、これはよくアクアショップでも聞く質問です。
CO2添加装置もなく砂底でよく増える水草をください、というような場合に…
こういうとき、よく候補に挙がるのがアナカリスことオオカナダモとマツモでしょう。
しかしながら、あちこちで帰化して大繁茂し、他の水草を駆逐している様子もしばしば見られます。同じくよく帰化しているコカナダモとの勢力争いでも、オオカナダモが有利に見える場合が多いです。
今回はそんな「金魚藻」の代名詞であるオオカナダモについて、ディープに語っていこうと思います。ほかの「金魚藻」であるカボンバ・カロリニアーナとマツモに関しても今後記事にしていくつもりなので、ご期待ください。
オオカナダモはすごい
劣悪な水槽環境でも育ちますし、底床がない環境に浮かべておくだけでも伸びていきます。分布域がラプラタ流域の亜熱帯域であるにもかかわらず、日本の厳しい冬も、酷暑に見舞われる夏も越すことができます。水温が35度近くなるような真夏のスイレン鉢ですら、オオカナダモは良好な生育を見せることがあります。ミズオオバコやスブタといった浅い湿地に生育する水草ではふつうのことですが、深い水に主に生育する水草としては驚くべきことです。
水槽内でも非常に高い適応性が見られます。CO2添加がなかったり、照明が弱い水槽でもオオカナダモは順応し、底砂がない水槽に浮かべておくだけでもしっかり成長できます。pH順応性も高く、他の水草がろくに生育できないような弱アルカリ性においてもオオカナダモは生育できます。なお、ソイル水槽などでは成長が非常に悪くなるのですが、これは特殊な条件と考えるべきでしょう。
他の水草が生育できないような異臭を放つ灰色の水域でも最後まで残っている水草はオオカナダモとコカナダモであることが多いです。水槽でも他の水草が育たない環境でも育つように、オオカナダモのサバイバル能力は突出しています。水流や温度の条件に関してもオオカナダモは非常に寛容です。まったくの止水域でも、高温になる浅い公園の池でも、深い池でも、濁っていても、澄んでいても、オオカナダモは水草が生育可能な場所に侵入すれば生き永らえるようにすら見えるほどです。
オオカナダモは極めてしぶといだけでなく、競争にも強い水草です。
オオカナダモは様々な水草にとって生育に適した湧水の影響がある水域においてはめっぽう競争に強いように見え、他の水草を駆逐して純群落になっていることがしばしばあります。コカナダモもしばしば侵略的になりますが、野外での様子を見ているぶんにはコカナダモとオオカナダモが競合したばあい、オオカナダモが優勢になることが多いです。いっぽうでコカナダモの方が拡散力が幾分強く、コカナダモが水源や細流を含めたどこにでも出現するのに対して、オオカナダモが侵入しているのは人口密度が比較的高い場所のことが多いように感じます。このことから、オオカナダモが侵入する原因は主に人為的なアクアリウムからの導入、コカナダモが侵入する原因は鳥による拡散や魚の放流の影響が大きいのではないか、と侵入状況を見ながら思うところです。
また新しい侵入地ができ、そこの水草が駆逐されないよう注意して扱わねばなりません。
ただ年々、どんな水草も生育できないような水域が大部分を占めてしまっているのが現状です。
個人的にはオオカナダモかコカナダモが生育できているようであれば、他の水草も何かしらが生存している可能性がある、と期待して見るようにしています。いまはそういう時代です。そしてそういう手掛かりをもとに、珍しい水草が見つかることもあります。
環境により可変する光合成システム
オオカナダモがきわめて頑丈な水草である理由は、この水草がきわめてフレキシブルに代謝を変化させ、環境に応じてその条件に最適なモードで生育するためであることがわかってきました。
オオカナダモの光合成は、CO2の不足する水中環境に非常によく適応しています。
基本的に、陸上植物は水中でCO2を使わざるを得ないものです。
藻類の場合はCO2を光エネルギーを用いて取り込んだりHCO3-輸送体があったりして、これらCCM(CO2 Concentrating Mechanism)により水中のCO2不足下でも無理やりCO2やHCO3-を詰め込み、HCO3-として蓄積したのちにこれをCO2に酵素を用いて変換します。しかしながら、陸上植物はより容易にCO2が手に入る空中の条件に適応してしまったためなのか、こうした機構を基本的には欠いています。
そのため水中に適応した水草といえどもCO2の吸収は拡散に頼ることになりますし、HCO3-を全くもって使えない水草も多いです。
オオカナダモの葉は透明で、切り飛ばすと水に沈みます。これは中に空気層がなく、葉が非常に薄いためです。
陸上に比べてCO2の水中での移動速度は一万倍遅く、陸上植物はそのため空中に葉をのばし、葉に多くのガスを含んで、葉の中でも局所的なCO2不足が起きないようにしています。しかし水中ではCO2の移動速度はどうすることもできず、止水条件において葉周囲および表面のCO2は絶対的に不足します。そのためオオカナダモは葉を薄くし、たった2層の細胞にしています。このことでCO2の拡散距離を短くすることができ、CO2の不足する水中でも光合成を効率的に行えるようになります。
しかしそれでも無理があり、葉表面のCO2は止水条件で活発に光合成が行われればいずれ枯渇してしまいます。光が強く、CO2濃度が低い場合は、オオカナダモは葉表面でCO2を生成する荒業にうって出ます。葉のおもて面に能動輸送でH+を汲みだし、上面のpHが低くなります。すると水中に(CO2よりは)ふつう豊富に含まれる重炭酸イオンを中和し、CO2が生じます。これを利用するというわけです。
さらに光条件が強くなり高温になると、さらに効率的なCO2供給が必要です。そんな場合、オオカナダモはふつうの陸上植物とは異なる、分類の難しいCO2濃縮機構を使い始めます。この機構はC4植物と同様の回路を用いますが、陸上植物の用いるC4回路が維管束鞘細胞を用いて行うのに対し、オオカナダモでは薄い葉細胞だけで完結させてしまいます。この濃縮機能により、オオカナダモは他の水草よりも高水温でもCO2を有効に用いて繁茂することができます。
オオカナダモは自身の代謝で生じたCO2も有効に活用しています。オオカナダモの茎は空気を多く含んでおり、水によく浮かび通気組織が発達しています。ここに自らの呼吸で生じたCO2を通気組織を介して茎に供給することによって、茎でも光合成をおこなっています。茎の含気により光合成の条件はむしろ陸上の葉と似ており、茎組織は陸上植物と同様に高いCO2濃度を要求する反面、CO2が効率的に供給されます。
オオカナダモは暗い条件にも非常に強く、100ルクスあれば生育可能といわれるほどです。さらに、光が弱い条件では茎が長く徒長することにより光を得やすい水面に素早く到着するようです。逆に、強すぎる光は苦手だともいわれています。
(執筆中…)
ギリシア人のみた水草たち (テオフラストス植物誌より)
今回は、人類の記録に残る最初期の水草記録についてみていきましょう。
自然科学の祖型は古代ギリシアに遡ることができます。より古い起源も辿れるかもしれませんが、少なくとも自然科学について深く調べ、考察し、文章を残し、そして今もなお読める、トレース可能な資料を探れといわれれば、やはり古代ギリシアといわざるをえないでしょう。
古代ギリシアにおいて、「植物学の祖」と呼ばれるのが今回紹介するテオプラストスΘεόφραστοςです。今回はテオプラストスの著書を和訳した「テオフラストス植物誌」大槻真一郎・月川和雄訳 を図書館で見かけたので、植物学のあけぼのの中で水草がどう認知されていたのかについて感想を書いてみます。
新訳版が後に出ているので、そちらも読まねば。
思ったよりはるかに面白いので、上下巻で1万円強する新訳版をカートに入れて本気で悩み中です。あとで図書館にないか再度探してみます。
この紀元前の著作を読んでいて驚かされるのがその観察力です。樹形、木の密度、根の生え方、木材の髄と外側の区別、ヤシ材と一般的な木材の違い、野生イチジクとそこに生じるイチジクコバチを用いた受粉法・・・などなど。流石に現在の視点からするとすべてが正しいというわけではなく、植物は種から発生するとしたうえで動物の自然発生説は認めるなどの時代的限界もありますが、その慧眼には驚かされるばかりです。
この書にはギリシア人の植物の育て方も載っています。紀元前の栽培法は是非実験してみたいところです。ロマンです。
テオプラストスは第四章「形態及び生息地の相違」にて、動物と同様に植物も陸上植物と水生植物に分けられる、とし、第14章「植物全体に関する、一般的な相違」において水生と陸生の区分を「最も特色があり、ある意味最も重要な区分」としています。テオプラストスが「水生植物」としたものは、湿り気のあるところでないと生育できない種や、湿り気のないところでは湿り気のあるところと同じ状態を保てず、質が悪くなる種です。これは現在いう水生植物に比べると非常に広い範囲で、沼地、湖、川、海、非常に湿った土地に生えるもの・・・つまり海藻と湿生・水生植物を含みます。たとえば、ヤナギやスズカケノキといった河畔林を構成する植物は水生植物であると考えたようです。しかしヤナギなどは水陸両生である・・・と書いていますし、ツルボランやナツメヤシについても陸上植物でありながらときに海に生えていると書いています。
これは水生といっていいのか?という悩みは当時もやはりあったようです。
これに限らず、彼は「自然はかならずこうと決まったものを持っていない・・・」そう幾つかのパートで書いています。
葉の構造について、多くの沼地の植物はカラモス(ヨシなどのこと)に似た、先のとがった葉をもち、葉は2つの部分からなって中間に竜骨状のものがある、としています。これは単子葉植物の披針形の葉のもつ主脈をあらわしていますが、これを竜骨にたとえるのは非常に理にかなっています。なぜならその支持する機能的にも竜骨と似たものがあるからです。沼地の植物、とされたのはおそらく、イネ科やカヤツリグサ科のことでしょう。
海藻および海草については、地中海(われわれの海)に小さなもの(読んでみると主に海草・海藻のこと)が、エリュトラ海(アラビア湾、ペルシア湾、紅海)に大きなもの(読んでみるとマングローブ植物のことです)があると書いています。
これらは4巻にあります。
イネに関しては4巻第四章、アジアに特有な樹木と草本(10)にあり、長い間水の中で育ち、殻をとると消化によい粥になる、としています。
4巻第6章では地中海の海草・海藻について述べています。当時の海草や海藻はピュコスとまとめられ、採集業者によって集められていたようです。(所謂本邦におけるモク採り)その中に様々なものがいることはわかっていました。さまざまな海藻や海草について述べられていますが、水草として特筆すべきは地中海を代表する海草、Posidonia oceanicaについて正確な記述がみられることです。広い葉で飾りひものような形状で、緑色で、根の外部は毛のようなもので覆われ、内部は幾つかの層からなり、根はかなり長くて太い…ということです。また、ギョウギシバに似たピュコスとして「ギョウギシバに似た葉をつけ、根もギョウギシバのように節があり長く横に伸び、カラモスのような茎を持ち、大きさは普通のピュコスよりずっと小さい」とあります。これは訳者によればベニアマモ科のCymodocea nodosaおよびアマモ Zostera marina、と書いてありますが、この記述はまさにZostera noltiiで間違いないでしょう!
大きさや形状、節があること(彼は底を非常に重視しました)からすると、コアマモ亜属のような小さなアマモ属か、ウミジグサを想起させるのですが、地中海にいるということはZ. noltiiしか候補に残りません。C. nodosaおよびZ. marinaは大きさがP. oceanicaにかなり似ており、ギリシア人をもってしても区別されていなかったのだろうと私は思います。
第7章のマングローブ植物の記述も素晴らしいものがあるのですが、私自身の知識量がまだ完全とはいいがたいので言及を避けます。それは第八章、「とくにエジプトの川、沼地、湖に生育する植物」に向けて余力を蓄えるためでもありますが…
第8章では様々な植物の名前が出てきますが、少なくとも旧訳版においては学名との対応関係がついていないものが多く、困ったものです。なので、なんとか読める範囲で解釈を試みようと思います。
パピルスが食用になるという話はヘロドトスやプリニウスも書いているのですが、テオプラストスもやはりそう書いています。食べ方は生で、また煮たり焼いたり液汁を吸いつくし、しゃぶりかすを捨てるというもので、両者のものと同じです。
気になるのが、根が木質が硬いので有用な炭になり、鍛冶屋が利用するという点です。今後観察するときに気にしてみようと思います。
ハスに関しては当時のエジプトにはありましたが、現在では見ることができません。エジプトのハスはペルシア経由で持ち込まれエジプトでは珍重されましたが、エジプト文明が崩壊したのち姿を消しました。根に棘があると書いているのは、葉柄基部のことでしょう。
スイレンは白花で日没とともに閉じ、日の出とともに開くとしています。これは奇妙なので、もしかすると時間の間違いかもしれません。果実が熟するとともに水中に潜っていくことについても(伝聞を含むためかかなり奇妙にですが)記述しています。食用とする部位は球根と種子としています。この球根は皮が黒く丸いとしているのでハスのレンコンではなく、スイレンの塊茎として間違いないでしょう。
トリボロス、はハマビシの属名にもなっている「棘のある植物」を指しますが、テオプラストスがいうトリボロス(第9章)は明らかにヒシのことをさしています。茎は先端に向けて太くなり、先端に長い柄をもつ幅広の葉と実をもち、茎は水底の根に至っており、茎からは毛状のものが多数出ている、実は黒く非常に硬い・・・まさにヒシでしょう。ヒシの水中根は現在でも混乱を生むものですが、テオプラストスも相当悩まされたようです。葉でもなく茎でもないので考察を要する、としています。また、ひとによっては一年草であるという、ということも、ギリシア人の観察眼の鋭さを思い起こさせるものです。
「シデ」はボイオティア地方のコパイス湖に生育していたと書かれていますが、おそらくセイヨウスイレンではないかと思います。葉と花が水面に浮かび、花はリンゴ大、白い花弁を持ち緑の葉(萼片)が4枚、キビ大の赤い種子がでてくる・・・まさにセイヨウスイレン以外にあり得ないでしょう。
テオプラストスがかなり興味を引いたのは意外にもガマで、「最も特異」とまで書いています。テオプラストスは地下茎について理解に苦しんだようで、主にこれに関して書いてあります。11章ではヨシ類と様々な利用、12章ではイグサ類について触れています。
とここまで読んでから巻末に対応表があったことに気づき、読み比べ…
まだいろいろ情報がありそうなので、調べ毎の合間にまた何回か読み返してみようと思います。
水草オタクの水草講座 マリモは水草なのか?
マリモは水草なのか?という質問をよくいただくので、この観点から水草とは何なのかについて紹介しましょう。
ごく最近ではマコモになってしまっているような気もしますが…
イネ、スイレン、ホテイアオイ、オオカナダモあたりではないでしょうか。
この4種を選んだのには理由があります。
水辺の植物は
①根本が水に漬かり、水の上に葉をのばす植物=抽水植物 (Emergent plantの訳語)
②草体の殆どが水に漬かり、水面に葉を浮かべる植物=浮葉植物(Rooted floating plantの訳語)
③草体全体が浮かんで育つ植物=浮遊植物(Free Floating plantの訳語)
④草体全体が水に沈んで育つ植物=沈水植物(Submerged plantの訳語)
の4つに分けられるからです。
それぞれの植物が、知れば知るほど水辺へうまく適応しており、水への適応度の高さという観点から安直に比較するのは難しいのですが、その中で最も水草らしい・・・水中への適応が特殊化した植物を挙げるとするならば、やはり④の沈水植物でしょうし、「水草」と聞いたとき、真っ先に思い浮かべられるのは沈水植物だと思います。
ここで、最も知名度の高い水草として「マリモ」を想像した人が結構いることでしょう。
マリモは水に沈んでいますが、沈水植物なのでしょうか・・・? 結論をいうと否です。
まず、沈水植物は水生植物のひとつあり、マリモは水生植物として数えられることはないからです。
水生植物というのはAquatic plantsもしくはWater plantsの訳語であって、これらの語は伝統的に、水に適応した種子植物、シダ植物、しばしばコケ植物を指してきました。ここで「しばしば」とぼかしたように、水生植物の定義というのは厳密なものではありませんが、これまで提唱されたほとんどの定義について共通するのは「水に適応した陸上植物を水生植物と呼ぶ」ということです。そして学術的な場合には、水草=水生植物を指します。
水草とは非常に難儀な、ひねくれた生き物です。
陸上植物は水中に生育する緑藻類から進化したものですが、水から上がる過程でその構造を極めて複雑なものとしました。コケ植物では、乾燥から守るためにクチクラ層を厚くし、組織内に空気を取り込み、ガスの出入口となる孔をあけ、乾燥に強い胞子をつけるようになりました。さらにシダ植物では、水を隅々まで分配し機械的強度をもたせるために維管束をこしらえ、ガスの出入り口を微調整しやすいように可変して動かせるようにし、種子植物では受精に水を頼らなくて済むようになりました。
これだけ色々こしらえておいて、また水に潜ってしまったのが水草です。
気孔は多くの種で退化し、組織内の空気も多くの種で失われ、維管束すらも退化的な種があります。種子および胞子は保たれていますが、その繁殖方法は極めて多様化していて、どの種も「結構無茶してるな・・・」と思わせてなりません。
どうです、ひねくれているでしょう?
クジラが魚ではない、というのと、マリモは水草ではない、というのはほとんど同じ意味です。水草は水から上陸した植物が水に潜ったもの、クジラは水から上陸した動物が水に潜ったもので、水草とクジラは非常に似ている面があるといえます。
ここまで話してもなかなかピンと来ない人が多いかもしれません。
マリモと水草を拡大して比較してみるとその差に納得しやすいかもしれません。
緑藻類に属するマリモは、その進化の過程で陸上を経験したことがありません。
マリモの構造は極めて単純で、その構造はまさに”毛玉”にたとえられるでしょう。その毛玉を顕微鏡で拡大してみると円柱状の細胞が直列に繋がり、その所々でまた円柱状の細胞が分岐するようにつく、という非常に単純なものです。マリモはこの毛玉がちぎれたり、鞭毛のついた細胞をつくって繁殖します。その構造は、繋がって大きな個体を形成しているとはいえ、目に見えない小さなプランクトン性の緑藻類、たとえばクロレラなどとほとんど変わりません。マリモを水草とするならば、クロレラは水草なのか?ボルボックスは?などといった問題も生じるでしょう。
昆布やワカメ、テングサなどもまた、陸上植物とはほとんど無縁な水中ネイティブな存在であり、姿はそっくりですが水草ではありません。サンゴも水草に似ていますがあれはクラゲに近い刺胞動物です。
というわけでマリモは水草ではない、というのがひとまずの結論となります。
ここからは脱線するので、初学者は読み飛ばしていただいた方が理解によいかと思います。
コラム① じゃあ水草コーナーでマリモが売ってるのは間違いなの?
じつは、水槽に使う水草の語義は学術的に言う水草=Aquatic plantとは異なっています。海外のアクアリウム向けの水草の本をよく見てみると、水槽に入れる意味での「水草」という語に対応する英語やドイツ語はAquarium plantおよびAquarien pflanzenであり、Aquatic plantとはしばしば分けて用いられています。
Aquatic plantとAquarium plantは非常に似た語ですが、微妙に違う点があります。それこそがマリモやアオミドロの扱いです。マリモもアオミドロも糸状の緑藻ですが、Aquatic plantはどちらも含まないのに対し、Aquarium plantはマリモをしばしば含みます。それは水槽にわざわざ彩りとして入れるものだからです。
いっぽうでアオミドロはといえば…わざわざ入れないし排除対象なのでAquarium plantにすらいれてもらえません。
何とも人の事情が絡む話です。こういうニュアンスの違いがあるので、私は水槽に入れる水草については「アクアリウムプラント」と音訳して使うことにしています。世界にはアクアリウムに向いていなくても素晴らしい水草が沢山あるし、ときには水草としての生態や適性をほとんど持っていないにも拘らず、問屋や販売店の都合で水槽に沈められてしまう植物があるからです。
アクアリウムにおいて水槽に沈めて育てられない植物を「あれは水草じゃない」というのは、水草栽培にある程度経験のある方なら見聞きしたことがある表現でしょう。では、それらは本当に「水草ではない」のでしょうか?たとえばイネは水に根元を浸して生育しますが、水中で生育はできない抽水植物なので、水槽に沈め続けると枯れてしまいます。しかしイネはAquatic plantです。
コラム②そっくりな水草と海藻、まとめる語はないの?
水草と海藻(主に褐藻類で、葉緑体の起源からことなる極めて異質な生き物)は見た目がそっくりなものがあります。とくに海においては、海水に適応した水草と、海の中で独自に大型化した褐藻類や紅藻類が”水中の森”を作り、生態系に非常に大きな貢献をしています。こうした大きな水中の植物は魚やエビも棲みかや産卵床として語る際、しばしばまとめたほうがいい場合があります。こういう場合、英語圏ではMacrophyteという語がつかわれます。このMacrophyteの訳語には「大型(水生)植物」というものがあてられがちですが、これは上記の水生植物と混同を招きあまり良い訳語とは思いません。ただ、むしろ逆の意味で考えると理解しやすいかもしれません。この「マクロ」植物/藻類という語は、目に見えない、「ミクロ」な様々な微細藻類 Microphyteの対義語です。つまり、目に見えるような藻類や植物はでっかい、マクロな存在であるということです。
古生物学における水草たち(古生代~中生代)
水草は陸上植物が水辺や水中など、水の深くかかわる環境に適応したものである。
これは原則中の原則である。
しかし、陸上植物においても被子植物に限って言えばいまだに、じつはその起源は水中や少なくとも水辺にあるのではないかという説があり、分子系統解析では否定されがちなもののあながち捨てきれるものではない。さらに、シダ植物や裸子植物に水生種がいまなお希少であることはその傍証かもしれない。今回は、化石記録における水生植物についてふれる。
水生植物は化石がそれほど多いわけではない。しかしその一方で新生代のフサモ属Myriophyllumやマツモ属Ceratophyllum、ヒシ属Trapaなどのように、化石記録がかなり多いものも含まれている。それは水草が生育する環境が水辺にあり、そして水辺の嫌気的条件がしばしば化石保存に適していることが関係する。しかしアクアリストならだれもがご存じのように水草は基本的に柔らかく、枯死すればすぐ溶けるように腐敗してしまうのもまた事実だ。そのため化石記録は基本的に断片的か、硬い種子などの部分のことが多く、全草が復元できるものはひじょうに少ない。さらにその同定が怪しいものもたくさんある。現在みられる数百属におよぶ水草がどのように水生適応を遂げてきたのか、その各々を自信をもって追いかけることは現状極めて難しいといえるし、昔にはこんな変な水草がいた!と、自信をもって提示できるほど素晴らしい化石記録は残念ながら殆ど皆無だ。
ただ、白亜紀の水草に関してはいくつかだけれど特筆すべきものがあるので、今後個別に記事を立てていきたい気はしている。新生代の水草は残念ながらかなりのものが現生種に繋がってしまうので、面白いものを見つけるのに苦戦している。
古生代の水草たち
0.断り書き・・・カンブリア紀のヒルムシロ属は流石にありえないという話・・・
陸上植物は水中に生育した緑藻類の一系統が古生代…おそらくオルドビス紀からシルル紀に上陸したものを起源としており、水草とされるグループは多系統でかつ、ヒルムシロ科はじめとしてその多くは中生代後半に出現した被子植物に属していることは2024年現在、ほぼすべての科学的知見が同意しているところである。
したがって本稿は、邦書(中には専門的な書物を含む)に散見されるカンブリア紀のヒルムシロ属花粉化石などを用いて陸上植物が水草に由来することを論じる目的では断じてないことを最初に断っておく。ヒルムシロ属の花粉がでたという話が何に由来するのか、具体的な初出の文献をみつけられていないが、このような説が出て割と広く受け入れられてしまった理由はいくつかあるだろう。まずヒルムシロ属はその非常に単純化した体制から、きわめて原始的な被子植物であると長らく考えられてきた。なかにはミズニラ類と並べて最も原始的な維管束植物と論ずることさえもあったという。そのため、ヒルムシロ属が最初の水草とする説があったとしても特に当時としては違和感がない。第二に、ヒルムシロ属の花粉は構造が単純かつ一見特徴的で、古生物学においてよく知られていることだ。ヒルムシロ属の花粉は極めて単純な構造で、以下のように古生物学では呼称される。「穴はなく25~35μmで外膜には細かい網目状模様がある」と。似たような形状のものが生じえないとは言えない単純な構造であり誤認することは十分あるだろう。
カンブリア紀のヒルムシロは現在の進化史や生命史を少し知っている人ならば必ずツッコミが入るレベルの荒唐無稽な話なので、もし調べるのに熱心な方が見かけたとしても忘れるようにした方がいいと断言する。
1.最初期の”半水生”陸上植物たちと最初の水草(?)Taenioclada decheniana
カンブリア紀後期からオルドビス紀にかけて、さきに上陸したのは動物なのか、植物なのか、はたまた菌類なのか、諸説ある。植物と断定できる化石記録として、最も古いレベルにあり、かつ詳しくその生育状況がわかっているのがデボン紀前期のライニーチャートRhynie Chartから知られるクックソニアCooksoniaやアグラオフィトンAglaophyton、リニアRhynia、ホルネオフィトン Horneophyton、ノシアNothia, Trichopherophyton, Ventaruraなどといったステム維管束植物で、これらは少なくとも湿地に生育していた。シルル紀~デボン紀からはほかにも様々な陸上植物が発見されている。Zosterophyllonなど、ほかにも現在や古生代以降の維管束植物の分類群のどれにも当てはまらない植物がみられる。便宜的かつ簡便にその特徴について記すが、各種の構造がその後詳しくわかるようになると、それぞれの種の所属に?がつくことが多くなり、はっきりと分類できない状況になっている。
リニア門Rhyniophyta・・・胞子嚢は頂生、顕著な紡錘形であり茎は突起を欠き裸。
ゾステロフィルム門Zosterophyllophyta・・・胞子嚢は軸に偏生、球形か腎臓形で茎はしばしば針状突起を伴う
トリメロフィトン門Trimerophytina・・・上記2つおよび現生の門にあてはまらない。PsilophytonやPerticaなど
これらの植物は気孔こそもっていたため空中でのガス交換はできても、維管束による水の吸い上げは未熟であった。これらの植物はすでにグロムス類の菌根菌と共生していたが根、葉、茎の区別はまだ不明瞭であり、そのような構造があっても現生植物とどのような対応関係にあるかもはなはだ疑わしい。これらの少なくとも一部は半水生とは言えるかもしれないが、水草であるかどうかに関しては議論を避ける。たとえば、ゾステロフィルム門ではないかといわれているThursophytonは湖成層から発見されており、水辺に生育していたと考えられている。しかし今の感覚でいえば水辺に生えているのは水草的だが、この時代はまだ陸上植物が水辺から逃れるほどの生理的特徴を備えていない時代、これらすべてを水草といってしまうのは乱暴ではなかろうか。
しかし、最初の水草はこの時代から発見されている。
私は最初の水草として、デボン紀前期のTaeniocrada dechenianaを挙げようと思う。Taeniocradaの表皮構造はよく保存されているが、植物体の少なくとも下面には気孔が見当たらず、茎というか二叉分岐を繰り返す草体は扁平でその側方に胞子嚢をつける。Taeniocradaはリニアよりは派生的と思われているが、そうだとすれば維管束植物の進化のきわめて早期において水中への再適応・・・最初の水草化が起こったことになる。これなら自信をもって水草といえるのではなかろうか。分類はリニア門といわれていたが胞子嚢は先端付近に側生するようで、現状よくわからない。
2.最初の小葉植物Lycophyte
ライニーチャートから見つかる植物の中でも、そこから1段階進んでおり、異彩を放つのがAsteroxylonである。この植物は根状の分岐を持っており、どうやら現在の小葉植物Lycophyte…ヒカゲノカズラやミズニラなどを含むグループの祖型であるらしい(注意;植物学上の小葉植物と、地質学上の小葉植物は意味がまったく異なるので、前者を意味したいときにはヒカゲノカズラ植物とか、Lycophyteなどと呼ぶことをおすすめする)。Atroxylonの維管束はまだ未発達で、茎のみしか涵養していなかった。のちのDrepanocladusやBaragwanathiaは葉まで維管束が通っており、デボン紀前期には小葉植物の基本的な構造が確立されたことがわかる。見た目はヒカゲノカズラそっくりでそうした類縁関係があると目されている。ただしいまのヒカゲノカズラと違って顕著な胞子嚢穂は形成しない。初期のヒカゲノカズラ類はしばしば水生ないし半水生として復元され、事実水辺に生育していたと思われる。ただし、水草として扱うには今一つたりないので、もっと水に適応していそうなミズニラ類とその巨大な近縁種についてみていこう。
3.ミズニラとリンボク類
ミズニラ類、リンボク(レピドデンドロン)Lepidodendronやシギラリア Sigillariaをはじめとするリンボク目Lepidodendrales、その小型版のようでより遅くまで生き残ったプレウロメイア目Pleuromeialesは互いに近縁で、デボン紀後期までには出現が確認できる。しかしその胞子記録はより古くから知られており、リニア門やゾステロフィルム門から直接進化してきたのではないかとする説もある。リンボク目はしばしば水中に根を張っていたと考えられており、水に適応した胞子分散生態を持っていたとも考えられているのでここでは水草として数えてみよう。リンボク目は高さ30m以上に達する植物だが、そのうち10mは直立した一本の塔のような構造となっている。木部や維管束の発達は極めて貧弱で、皮層が充実することによりその径を増した、あるいは径を増さずに太くなってから直接伸びていったと考えられる。根はStigmarian root complexとよばれる。太い二又分岐する根のような構造から、繊細なに叉分岐する細根が無数に生じることにより根を張った。この太い根状の突出物はミズニラ属における茎基部の隆起、細根はミズニラ属の根と構造が酷似し、それぞれ相同であると考えられている。10m以上電柱のようにまっすぐ伸びた茎は先端付近で急に二叉分岐をはじめ、その先端に胞子嚢穂をつけた。この胞子嚢穂は水に浮かぶようにできていたと考えられており、水で満たされた根本に落ちたのちに水に浮かんで漂流分散し、泥底で発芽したと考えられている。上記からすると生育の最初期は沈水葉で育ったかもしれないと思わせるが、残念ながらその証拠は未だに見つかっていないと思う(知る限り)。リンボク目はデボン紀後期に出現し石炭紀に大繁栄したが、石炭期末には早々に絶滅してしまう。リンボク類をはじめとした初期の水辺の森林は岩石の風化を促進し、光合成による消費だけでなく水中への二酸化炭素溶解を早めて大気中の二酸化炭素濃度を効果的に下げたとする説もある。
プレウロメイアや”茎の伸長する”ミズニラ類は中生代まで生き残ったが、中生代の中ごろにはミズニラ類しか残っていなかった。そのミズニラ類も白亜紀末の大量絶滅で壊滅的な被害を受けたようで、現在世界に生育するミズニラ類は新生代にたった一種を由来とするものらしい。そのため現生のミズニラ類はふつう水生だが、中生代のミズニラ類が水の周囲や湿った場所に生えていたことはわかっても、ほんとうに水中に生えていたかは知る限りいまいちはっきりしない。気孔などの観察で分かることと思うので、今後の研究に期待したい。
4.謎に包まれた大葉植物の起源
ここまで話してきた小葉植物・・・ミズニラ類、イワヒバ類、ヒカゲノカズラ類は、その茎も根も頂点での二叉分岐しかせず、現存する他の維管束植物とは根も葉も相同でない可能性が示唆されてきている。植物体の構造はほかの陸生植物とは大きく異なり、それらは収斂進化の可能性が高い。したがって、現在大部分を占める大葉植物の祖先形がどのようなものであったのかは闇の中に包まれている。わかっていることは、デボン紀後期の時点で種子植物的な特徴を一部持つ前裸子植物まで様々なグループが爆発的に出現していたことだ。
5.トクサ類の登場と水辺での長きにわたる繁栄
トクサ目はデボン紀後期には出現した。最初期からその姿をあまり変えていない維管束植物である。最初期のトクサ類であるプセウドボルニアPseudoborniaやアルカエオカラミテス科Archaeocalamitaceaeはすでに巨大な植物で、10~20mもあったと考えられている。石炭紀の森林でかなり大きな位置を占めたロボク Calamitesは現在のスギナを高さ20mにしたような滑稽な姿をしている。その枝は針状で、分離した単体ではアニュラリアAnnulariaとよばれる。ロボクも湿生植物の傾向がつよく、水辺でのみ生育したことがわかっている。小型だが異彩を放つデボン紀~石炭紀~ペルム紀のトクサ類がスフェノフィルムSphenophyllumだ。このトクサ類は現生種とは異なり、幅の広い葉をもっていて、つる性植物だったとも考えられている。石炭紀の時点で現在のトクサそっくりなトクサ類も出現していた。古生代に隆盛を極めたトクサ目だが中生代に入るとトクサ類しか残らず、ペルム紀~三畳紀の抽水植物であるネオカラミテスNeocalamitesに代表される大型化した系統もあったが中生代中期の乾燥化によりトクサ類の分布は水辺に局限されるようになった。水辺においてトクサ類は中生代を通じて一般的なグループであるが、これも新生代に至るまでの間に大打撃を受けたらしい。現生トクサ類もまたミズニラ類と同様に、新生代にたった1種から分岐したものと考えられている。
6.デボン紀後期の水辺の森林
デボン紀後期の水辺の森林を構成したリンボク類、トクサ類を紹介したが、原裸子植物についても軽く触れる。アルカエオプテリスArchaeopterisは水辺に巨大な森林を作った初期の巨大植物であり、NHKスペシャル「地球大進化」でも大きく取り上げられたので知っている人が多いかもしれない。この植物は一見シダのような葉を持っているが、その小葉のようにみえる一枚一枚が葉であるという複雑な枝系だったと考えられている。維管束の構造も裸子植物的である。他にも様々な裸子植物的な形質を持つ維管束植物がデボン紀後期から発見されており、その時点で裸子植物の祖先にあたる系統がすでにあったと考えられる。これらは原裸子植物と呼ばれている。
真正のシダ植物の古生代における化石記録もまた多いのだが、明らかに水生といえるようなものは知らないので割愛する。水生シダ植物は中生代の項で解説する。
7.シダ種子植物と最初期の針葉樹、そして最初のマングローブ
古生代からはさまざまなシダ様の植物が見つかるが、その多くがシダ植物とはいいがたい存在であることがわかってきている。これらはしばしば裸子植物的な巨大な”種子”を形成し、マクロな構造的にも、ミクロな構造的にも花のような構造と花粉のような胞子をもっていた。アレトプテリスAlethopterisなどが代表的だろう。こうしたもののなかで、メデュロサMedullosaは淡水からわずかに塩分の入る湿地で生育したと考えられている、木性シダ様の抽水になりうる植物だった。シダ種子植物、ソテツ類、ベネチテス類、針葉樹類、グネツム類は裸子植物を構成したと考えられる。最初期の針葉樹類としてコルダイテスCordaitesがあげられる。この種はマングローブを構成する植物であったようで、現在のヒルギ類に似た気根を形成し海岸付近の若干ながら塩分の混じる水域に群生していたと考えられている。
中生代の水生植物
1.中生代の植生
中生代の植生はペルム紀迄の影響を強く受け、乾燥化が進んでいくもののある程度の湿度が保たれシダ植物が反映した三畳紀~ジュラ紀中期、乾燥化が進んでいき乾燥耐性の強いケイロレピディア科Cheirolepidiaceae裸子植物やソテツ植物、一部のベネチテス類が優占する熱帯~亜熱帯と湿潤かつ温暖なシダ植物や落葉性裸子植物が優占したジュラ紀後期~白亜紀前期、爆発的に被子植物が増加して裸子植物の多くを駆逐し、熱帯雨林ができはじめたりするなど現在の気候や植生ができあがっていった白亜紀後期、にわけられるだろう。水草は前半において非常に少ない。特筆すべきは白亜紀前期のことであり、被子植物の多様化も水辺で始まった。ただし、古生代後半と比べて概して化石的証拠が少なくぱっとしないのが中生代であり、巨大な植物食動物の知名度とくらべて当時の植物には未知の部分が非常に大きい。植物体全体を復元できるものすら、ごく僅かしかおらず、中生代の植生は実質的に絵に描くことができない。白亜紀前期までの中生代を通じて支配的であったケイロレピディア科やベネチテス目、木本性のですら満足のできる復元ができるものは皆無であるといってよく、葉と幹の組み合わせがわかるものは数種だけだ。殆どの復元はバラバラになったパーツ群をありそうな、見つかっている組み合わせでなんとか無理くりつなぎ合わせたものでしかなく、その枝ぶりや幹と葉の組み合わせは多くが想像に頼っている。
2.トクサ類とシダ植物、あとコケ植物
中生代を通じてトクサ類は水辺に豊富である。ミズニラ類も多く見つかり、化石こそ少ないもののヒカゲノカズラ類の胞子化石は豊富である。
いまいるようなシダ植物は石炭紀にはシダ種子植物に埋もれていたが確かに存在していた。中生代の時点で広義ウラボシ目を除くグループがだいたい出現しており、ゼンマイ科、フサシダ科、ウラジロ科、マトニア科、ヘゴ科、ヤブレガサウラボシ科などが湿った地域に繁栄していた。ディクソニアは中生代からも知られるが、ジュラ紀後期から白亜紀前期の種は水辺に見られたようで、また必ずしも現在のように木性ではなかった。
デンジソウ科はRegnellitesに代表されるように白亜紀前期頃から少なくとも見つかるようになる。中生代のデンジソウ類はRegnellidiumに似た2枚葉だった。サンショウモ属やアカウキクサ属も白亜紀には出現している。しかしそれも白亜紀のこと。それまで水辺にいる維管束植物といえば抽水性のトクサ類やミズニラ類程度だったと思われる。
コケ植物の化石記録は少ないが、三畳紀のNaiadita lanceolataは水生コケ植物である。
3.裸子植物のマングローブ?
シダ種子植物は中生代になるとめっきり少なくなり、カイトニア目Caytoniales、コリストスペルマ目Corystospermales、ペルタスペルマ目Peltaspermalesしかのこらなかった。コリストスペルマ目のPachypteris papulosaはその分厚い葉から塩性湿地に生えるマングローブ植物だったと考えられており、二分岐を繰り返す枝に長大な複葉をつけた。さらにこの植物はおそらくケイロレピディア科針葉樹のBrachyphyllum crucisとともに見つかることから、混生して裸子植物のマングローブ林を作っていたかもしれない。裸子植物の塩性湿地への適応としてはほかに、虫媒も示唆される特殊な針葉樹であるケイロレピディア科のFrenelopsis ramosissimaが取り上げられる。しかしこれに関しては否定的証拠もある。厚い多肉質の葉は塩性湿地だけでなく乾燥への適応でもあり、沿岸地帯はしばしば乾燥することに注意は必要だろう。
いっぽうで、裸子植物は知られる限り水中には進出せず、中生代の長い間の水中は古生代と変わらず、シャジクモ類が支配的だったとみられる。しかも卵胞子以外の化石記録がろくにない。(卵胞子は豊富で、時代を決めるのに使われるくらいだ(示準化石))
4.白亜紀前期、突然出現し陸上に先行して広がった沢山の水草たち
分子系統的に被子植物の水生起源説は否定されがちだが、しかし化石記録からは初期の被子植物が水辺からまず分布を広げていったことは明らかである。最初に出現する被子植物はバレミアンのスペインから産出した根を欠く沈水植物であるモンセキアMontsechia vidalii(マツモ科?)で、次もバレミアンの中国から産出し、水生植物が示唆されるアルカエオフルクトゥスArchaeofructus、そしてそれに続くアプチアン期にはニムパエイテスNymphaeites、ネルンボNelumbo(ハス科)、”Ranunculus”(キンポウゲ科??*R. ferreriはバイカモ様)、Protaephyllum(分類不詳)、Hydrocotylophyllum(分類不詳)、ノトキアムスNotocyamus(ハス科)、ジャグアリバJaguariba(スイレン科)、オロンチオフィルムOrontiophyllum(サトイモ科?)、Plantaginopsis(分類不詳)Alismaphyllum(分類不詳)Vitiphyllum(分類不詳)、Menispermites(不詳)、Spixiarum(サトイモ科?)、Pseudoasterophyllites(マツモ科?)、Pluricarpellatia(ハゴロモモ科)・・・と、水草ばかりといっても過言ではない。水辺の植生は最初に被子植物によって占領されたと考えられている。そして陸上においては、水辺にやや遅れて被子植物が増えてくる。そして、この爆発的な多様化イベント以前からは被子植物の化石が一切出現しない。これに関しては、初期の被子植物が洪水直後のような撹乱環境を好んでいたため、そうした環境に生育する湿生・水生植物から多様化が始まったとする意見がある。同様のニッチを狙う戦略をグネツム類もとったようで、嫌気性の他の植物がみられない沼地から多くのグネツム類が見つかっている。そうしたニッチからして潜在的には、水草としてのグネツム類が将来的に見つかるポテンシャルがかなりある。マオウやウェルウィッチアなどのグネツム類の多様化も、被子植物とほぼ同じか、それに僅かに先行して行われていたが、被子植物に負けてしまったものと思われる。ただし白亜紀の一部の「水草」がじつは水生グネツム類ではないかと書いている文献もあるため、断定的なことは非常に言いにくい。奇妙なことだが、シダ植物のサンショウモ科やデンジソウ科の水への進出も同じ時期である。
短いターンオーバーが可能な草本であったためそれまでの木本やシダ植物などと違い短期間で殖えられたのが被子植物の利点ではないかとする意見があり、白亜紀中期の極端な気温上昇とCO2上昇(最大で1800ppmに達し、これは人間が二酸化炭素中毒で眠気を催すレベルを超えてしまった)によっておきた様々な天変地異による既存の植生破壊や、同時期に出現した鳥類(アプチアンの時点で水かきをもつ水鳥が多かったことが足跡化石からわかっている)の爆発的放散は被子植物に有利に働いたことだろう。
白亜紀後期には今見られるような水草の一部がすでに見られていた。スイレン科、ハス科、ガマ科、サトイモ科(Orontium)などが挙げられる。もちろん今では絶滅したグループもあり、QuereuxiaやHaemanthophyllum、Limnobiophyllum(サトイモ科)、Cobbania(サトイモ科)、水生シダでサンショウモとシダをつなげたようなHydropterisなどが挙げられる。陸上植物も被子植物が支配的になっており、水中も陸上も現在につながる植生の基礎は白亜紀中期にできたと考えられている。しかしながら白亜紀は化石が保存されにくい時代で産出状況がバレミアン―アプチアン以降かなり悪く、次に非常によい化石が見つかるようになるのは鮮新世になってからのことである。そのためその間に具体的にどんなことがあったのか、よく言われる昆虫の多様化との関連などは謎が多い。なお私の観測では昆虫の多様化の方が先行しているような気がしてならないのだが…
***************************************
新生代の水草に関しては現在につながるグループの話が多いので一旦後にします。
ただ、アゾライベントについては触れておいた方がいいかもしれませんので、各自検索して自習してください。
水草オタクのレンコンがたり.
寒くなってきました。
今回は、正月に欠かせない冬の作物、レンコンについて、水草オタクがみっちり語っていこうと思います。
- I. 世界のハス
- II. ハスの進化史と分類的位置
- III. ハスの草体について
- IV. レンコンの穴とハスの通気システム
- V. 葉による加圧と気体循環の制御
- VI. まだわからない部分もある、レンコンの構造
I. 世界のハス
レンコンはハスの肥大した地下茎です。では、ハスの仲間は世界に何種あるかと言えば、2種しかいないと2024年現在では考えられています。アジアからオーストラリアに分布するハスNelumbo nuciferaと、北米(~南米北部?)に分布するキバナハスNelumbo luteaです。キバナハスは花が黄色であるだけでなく、草体は概して通常のハスよりは小柄で葉柄に棘がなく、花弁数が少なく葉色が濃い点で区別できます。栽培に関して言えば、キバナハスの方が難しい傾向があるようです。ハスとキバナハスは古生物学的にはかなり昔に分岐したと考えられてきましたが、分子生物学的には更新世ごろ、もしくはそれ以降に分岐したもののようです。
ハスはインドおよび中国において古くから利用され、東洋のイメージが強い植物といえます。ハスとスイレンを区別する際、しばしばハスには”Indian Sacred lotus"であるとか”Indian bean"、”Chinese water lily"と呼ばれるのはその傍証でしょう。ヨーロッパにハス類の分布はなく、Nelumboという属名はシンハラ語に由来するといわれています。
アジアにおけるハスの利用および改良は、どうも中国のものと南~東南アジアのものに分けて考えた方がよいように思われます。アジアのハスは温帯性ハスと熱帯性ハスの2系統に分けられ、今回紹介するレンコンがとれるのは温帯性ハスのみです。
熱帯性ハスは細いランナー状の根茎が主体で冬場の根茎の肥大は顕著でなく、同じ条件で育てた場合には熱帯性ハスのほうが2~3か月長い開花期間を持ち、常緑性の傾向があります。国内で見かけることは殆どありませんが、タイやベトナムでは肥大しない根茎を食用にします。日本においてもベトナム料理店には稀ながらも熱帯性ハスの根茎が売られています。熱帯のハスはレンコンを作らないことや、またインド(特に南部)では神聖視されて食べない、仏教の影響が強い地域でも神聖視される面が強い・・・などから、食用としての改良は南・東南アジアでは盛んではないようです。そのため研究が遅れており、どのあたりの地域まで熱帯性ハスがいるのか、熱帯性ハスの中での遺伝的分化はどの程度進んでいるのか、興味深いところです。
温帯性ハスは根茎用ハス、種子用ハス、花ハスの3つに分けられることが多いですが、それらはしばしば多系統であるようです。花ハスはおもに中国で発展した栽培品で、草体は小型ながらも花が高くあがります。種子利用ハスの分類ははっきりしませんが、(多系統ともされる)一般に栽培の中心は根茎利用ハスより南方に位置します(湖南省、江西省、福建省、ベトナム)。これらの種子利用ハスは”亜熱帯性の”分布を示すように思われますが、今後の研究に期待したいところですし、もし可能であれば各品種を実際に栽培して形質を比較したいところです。
今回の主役である根茎利用ハスもまた(日本で利用されるものを含めて)中国由来の品種であり、一般的には白花で開花数は少なく、草体は大型である傾向があります。根茎利用ハスは日本においても中国においても最も作付け面積の広いハスであり、レンコンは日本、中国ともに一般的な食材です。中国においては湖北省、江蘇省、安徽省、浙江省で栽培されており、日本では茨城県が生産量の半数近くを占めています。
日本で育てられている花ハスおよびレンコン用の食用蓮、繊維用ハスはすべて温帯性ハスおよび、温帯性ハスとキバナハスの交雑に由来します(すくなくとも温帯性ハスとキバナハスは交配可能で交配種にも稔性があります)。さらに、食用ハス、花ハスともに主要な品種は中国に由来し、日本由来のハスはごく一部の”古代ハス”と呼ばれるような個体群などを除けばほぼ栽培されていません。
西洋およびオリエントにおけるハスの存在感はほとんどないのですが、エジプトではかつて珍重された経緯があります。スイレン類(Nymphaea nouchali var. caeruleaとNymphaea lotus)が古典的に利用されていたエジプトにハスを持ち込んだのはおそらくアケメネス朝ペルシア帝国で、前5世紀ごろから用いられるようになったようです。エジプトのハスは自然分布ではありませんが、エジプトにハスが持ち込まれるとイシス教において神聖視され、古来より用いられてきたスイレン類にとってかわったようです。但し、エジプトに移植されたハスは現存しておらず、スイレンと似る点も多いことから記述および当時の絵に頼るほかありません。
II. ハスの進化史と分類的位置
ハスは生きた化石と言えます。双子葉植物のなかでも特に原始的な植物であり、ハス科植物はハス属以外の属は現存していません。ハス科は真正双子葉植物の中でも最も早期に分岐したヤマモガシ目に属しますが、ほかのヤマモガシ目植物とは白亜紀前期の時点ですでに分かれていたと考えられています。最近記載されたNotocyamus hydrophobusは1億2100万年前の白亜紀アプチアンから産出しており、最初のハス科植物ながら白亜紀のハス科植物としては全草に近い保存をされており、現在のハスのものに似た花托がみられ、すでに水生であったと考えられています。ハス属とされる化石は約1億1000万年前のアルビアンから産出するN. lusitanicaと N. weymouthiが最初で、双子葉植物の属としては最も古い部類に入ります。白亜紀からは知る限りで5属22種のハス科植物が知られており、ハス属としてはN. choffati, N. kempii, N. intermedia, N. primaeva, N. laraimiensis, N. tenuifolia, N. lakesiana, N. crossii, N. dawsoni, N. puertae, N. provinciale, N. arctica, N. amurensis, N. orientalis, N. schweinfurthi、ハス属以外としてはNelumbites virginiensis、Paleonelumbo macroloba, Nelumbago peltata, Exnelumbites callejasiae, Notocyamus hydrophobusが挙げられます。これらの多くは葉のみから記載されていますが、現在のハスと比較すると葉中央の円盤状部分を欠いていたり(このことはのちに述べるが生理的に極めて重要な点と言える)、葉に鋸歯を持っていたりするなど、現在のハスと祖先を同じくするものの、現在のハスの祖先というわけではないものが多いようです。ハス科植物は白亜紀の時点では少なくとも北米、ユーラシア、アフリカ、南米に分布し、ほぼ世界中と言っていい分布を確立したと考えられます。ハス科植物の繁栄は隕石衝突による白亜紀末の大量絶滅後も(少なくとも北半球では)続き、N. protolutea, N. aureavallis, N. changchangensis, N. nagalensis, N. nipponica, N. weylandi, N. protospeciosa, N. endoana, N. hungarica, N. lacunosa, N. megalopolitana, N. minimaなどたくさんの種が記載されています。しかし第四期の氷河期により分布は著しく縮小し、アジアと北米にしか生き残れなかったと考えられています。ハスは寒さにそこまで強い植物ではなく、レンコンの保存適温も5度以上、5~8度が適温とされています。
III. ハスの草体について
ハスの草体は横走する地下茎(肥大化したものが所謂レンコン)と、その節から出る根、葉、花茎から構成されます。葉は各節から1枚ずつでるように見えますが、基部には発達しない、葉鞘状の低出葉Cataphyllが2枚あるため、各節から出る葉は3枚ずつであると考えられています。葉は盾状(Peltate)と表現される、長い葉柄が円形ないしやや漏斗状になった葉の中央下部につく形態をしており、葉柄には棘があります。(キバナハスにはない。)葉柄の断面には4つの大きな穴および、5つの小さな穴があり、これらはGas Canalと呼ばれます。ハスは地下茎の節から発根し、”レンコン”の側面にあたる節間からは殆ど発根しません。地下茎の節には3つの小室があり、節間にあたる”レンコン”にもGas canal・・・すなわち穴が約10個あります。花は1つずつつく両性花で、放射相称性を示します。花は直径10~30㎝ほどのものが多く、ハスの場合白色~桃色です。花は水面より高くつくものの、葉柄と花柄はほぼ同長か花柄の方が短いことが多く、葉より下に花がみられることもしばしばです。花の萼片や花弁の数は可変が激しく、萼片数は2~5、花弁数は20~30、雄蕊数は200~300とされます。雌蕊は20~30心皮で大型化した花托に埋もれます。この花は発熱することが知られており、また雌蕊の構造も興味深いのですが今回はレンコンの話なのでこの辺りにしておきましょう。
IV. レンコンの穴とハスの通気システム
ここからが本題です。
レンコンの穴はしばしば「通気のため」とされ、また葉柄の穴に関しても水を通すハスの葉シャワーや夏休みの自由研究などの題材にしばしばされてきました。しかしながらレンコンの穴とそれを介したハスの通気システムは植物の中でも極めて特殊かつ複雑な構造であり、他の水生植物とは一線を画する面があります。この構造全体の把握はとても難しく、100年ほどにわたって研究されている状況です。ここからは、レンコンの穴を通じて、1億年かけて完成されたハスの”異常な”水生適応について書いていきます。
レンコンの穴は一見放射相称に見えますが、じつは上下軸が存在する左右対称な構造です。しかも、そのそれぞれの穴には特有の機能と複雑な接続関係があり、同じ穴が並んでいるだけではないのです。
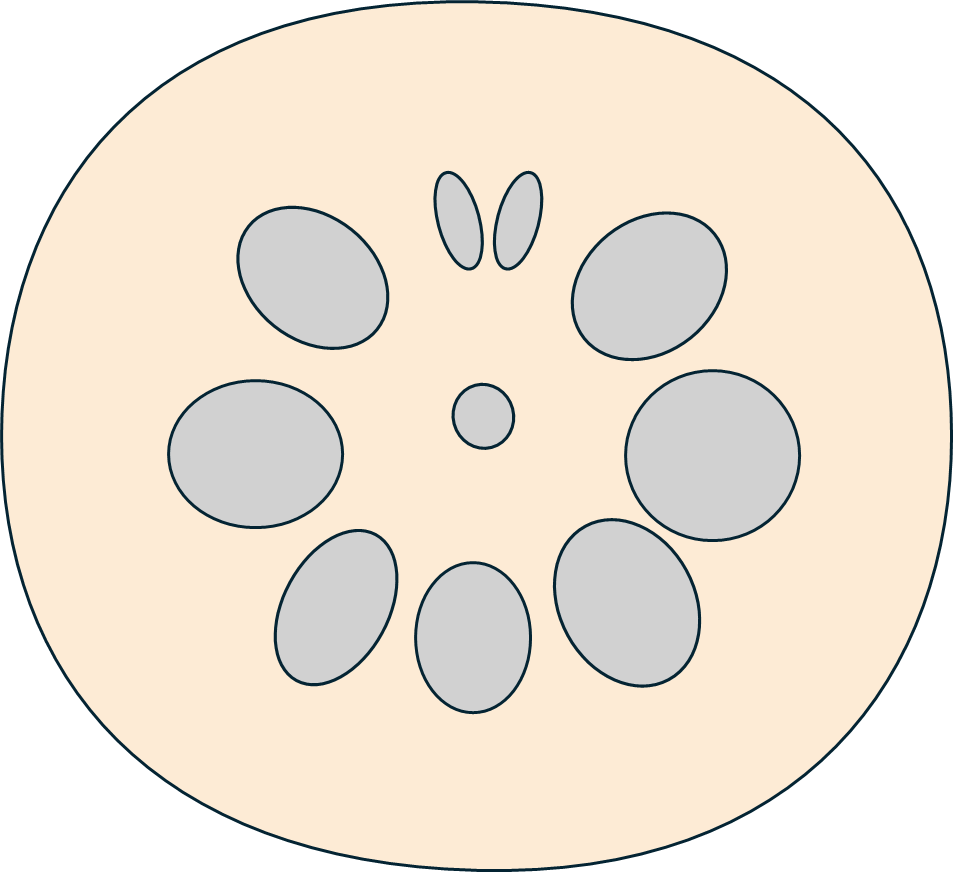
レンコンの穴がだいたい10個であることはよく知られていますが、それに上下軸があることはほとんど触れられていません。レンコンの大きな穴はだいたい10個ですが、その中にはほぼ必ず、小さい1ペアとその対極に位置する大きな1穴があります。それが軸になり、小さい1ペアが上側に位置します。
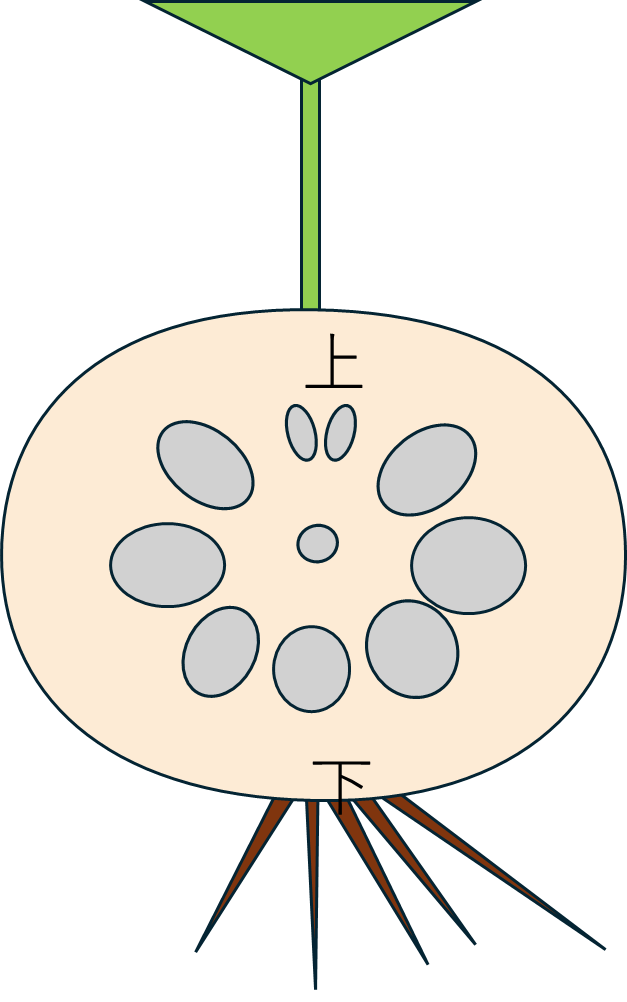
断面側から草体をみると、こんな感じになります。
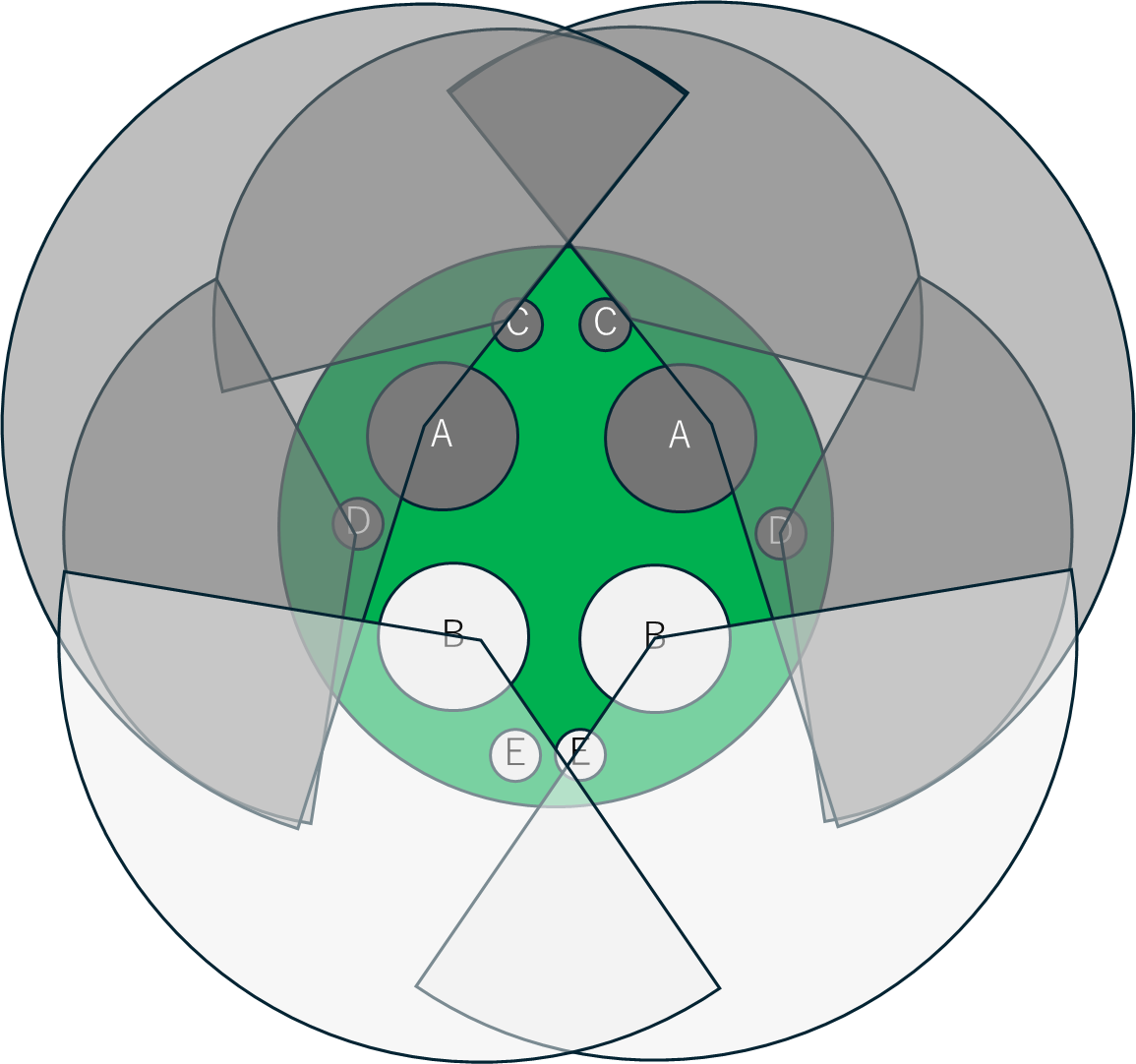
次に、この穴に名前を付けます。

次に、ハスの茎の穴にも名前を付けます。ハスの茎の穴もやはり放射相称ではなく明確な極性があります。

このなかで、BおよびEで示した部分からは水中で切っても気泡が出ず、A, C, Dから気泡が出ることが知られています。つまり、A, C, Dは根茎からの上向きの流れ、B、Eは葉から根茎への下向きの流れを担当しています。ハスの通気システムは基本的に各穴を流れる気体の流れは一方向性かつ規則的で、ほかの水生植物に見られるものとは明らかに異なっています。
これらの穴が節においてどのように接続しているかについて模式的に図示します。レンコンの節はなかなか観察しにくく、シリコン型をとって研究が行われています。節には3つの小室があり、X, Y, Zで示しています。

Matthews, P. G., & Seymour, R. S. (2006)より作図。
葉から送られてきた空気は小室Zを通ってレンコンの側方2対にあたるIIおよびIIIを通って草体の前後に配分されます。IIを通る空気の一部は小室Xに接続して上側に位置するIに入り、Iはさらに下流側で小室Yを介して葉柄のA, C, Dに接続し葉に送られます。
きわめてややこしいので、1方向にするとこのような図になります。

ハスの葉柄はガスの移動する極性が1本の葉柄の中でも分業していることが知られていますが、こうしてみると基本的には新しい葉から地下茎を2節通って古い葉に流しているように見えます。水中に空気を効率的に送るには、入り口と出口がある一方向性の経路が必要です。特に大型化する水生植物、とくに深い水から抽水・浮葉するためにはその空気輸送距離はそうとうに長いものになり、効率的な輸送システムが必要となります。ただ穴をあけているだけでは不十分なのです。他の浮葉性水草(たとえばコウホネやスイレン、アサザなど)では、新しい葉から入ったガスを地下茎に通し、古い葉から排出するシステムが用いられていますが、基本的にはこれに似た、しかしより複雑なシステムといえます。
しかし、それで終わらないのがハスです。
レンコンの謎が少し解けましたが、その先を見るにはハスの葉に目を向けねばなりません。
V. 葉による加圧と気体循環の制御
ハスの葉の中心に溜まった水から、ブクブクと気泡が出てくることはかなり前から知られてきましたし、気にしてみるとしばしば見かける光景でもあります。
ハスの葉の中央部には白っぽい円盤状の部分があり、これが地下茎から葉柄を伝って出てきた空気の出口となっています。この部分(Central plate)には特殊な大型の気孔があり、根茎からでたガスの出口となっています。ハスのCentral plateの気孔は午前中に開き、正午に閉じ、午後に再び開くという周期的な動きをしており、草体内のガス需要や、また草体から流出するガス流量、ないし草体内で生じたCO2の光合成への利用が制御されていると見られます。
Central discに接続するのは3系統ある葉柄を上行するGas canalのうち最も太いAの一部のみで、残りは一旦集まったのちに葉脈に沿って広がります。逆に、葉脈に沿った通道組織はBとEにもまとめられ、葉柄を下行します。傘状の葉にどのGas canalがつながるかは葉の部位によっても異なります。概して、葉の2/3が排出につながるGas canalに、1/3が空気の取り込みにつながるGas canalに向かうようです。

Vogel, S. (2004).より作図。
Vogel, S. (2004).を参考に作図。
さて、ここまでハスの発達した”一方向性の”ガス循環についてみてきましたが、その駆動力についても触れてみます。
ハスの植物体内を通るガスは光合成ガスではなく、大気中の組成とほぼ変わらないガスが送られていることがわかっています。
ハスの植物体内へのガス取り込みは気孔を介しており、ハスの気孔はまるで浮葉植物のように上面のみに分布し下部や葉柄、花柄にはほぼ欠落しています。(スイレンと違って抽水植物なのでハスの葉には下部にも気孔がある、とする誤解が散見されるので注意。)気孔は完全に閉じることができず、常にある程度開口しています。
気孔から入った空気がハスの葉で加圧され、日光による熱によりその加圧が推進される(Thermo-osmosis)ことはよく知られていますが、これはあくまでも加熱により加圧が起きるのであって、光合成による影響ではないことがわかっています。ハスの葉の索状組織の下には密な柔組織があり、その下に空気の入った小室がみられます。小室は葉の断面の半分以上を占め、小室内で蒸発した水分による葉内外の水蒸気差によって外気の乾燥した空気を取り込む(hygrometric pressurization)と考えられています。他にも、温度差があり半透膜を隔てたガスが半透膜を通って移動するKnudsen効果もその駆動力として考えられることもあります。加圧された空気は柔組織により外に逃げることが防がれ、小室につながるGas canalへと導かれてガス循環の駆動力となるようです。
VI. まだわからない部分もある、レンコンの構造
今回の記事を書くにあたってハスの構造を復習していたところ、ハスの葉柄をフィルターとして使えるという自由研究が出てきて驚かされました。
ハスの葉柄の穴にはフィルター状ないし網目状の水を弾く膜があり、この構造の意義に関してはなかなか解釈の一致を見ないところです。一つの説としては破損した茎から水が侵入するのを防ぐというものがあり、たしかに実験上は機能するようではあるそうです。同様のネット状ないしスポンジ状の構造は根茎の節部にも存在し、計算上は同様に水が地下茎に侵入することを防げるとのことですが、他にも肺のように酸素を取り込む機能など、ほかの意義がある可能性もあります。
食卓のレンコンにはまだまだ謎が沢山ありそうです。
Prigioniero, A., Scarano, P., Ruggieri, V., Marziano, M., Tartaglia, M., Sciarrillo, R., & Guarino, C. (2020). Plants named “Lotus” in antiquity: Historiography, biogeography, and ethnobotany. Harvard Papers in Botany, 25(1), 59-71.
Mekbib, Y., Huang, S. X., Ngarega, B. K., Li, Z. Z., Shi, T., Ou, K. F., ... & Yang, X. Y. (2020). The level of genetic diversity and differentiation of tropical lotus, Nelumbo nucifera Gaertn.(Nelumbonaceae) from Australia, India, and Thailand. Botanical studies, 61, 1-11.
Tian, H. L., Xue, J. H., Wen, J., Mitchell, G., & Zhou, S. L. (2008). Genetic diversity and relationships of lotus (Nelumbo) cultivars based on allozyme and ISSR markers. Scientia Horticulturae, 116(4), 421-429.
Zhou, P., Jin, Q., Qian, P., Wang, Y., Wang, X., Jiang, H., ... & Xu, Y. (2022). Genetic resources of lotus (Nelumbo) and their improvement. Ornamental Plant Research, 2(1), 1-16.
霞正一. (2003). 食用ハスの交配育種法: 遺伝資源の収集から品種登録まで.
篠原啓子, 圓藤勝義, 澤田英司, 永峰賢一, 水口博之, & 福井裕行. (2016). 食用ハス (Nelumbo nucifera) における肥大根茎 (レンコン) の形状の品種・系統間差.
Gobo, W. V., Kunzmann, L., Iannuzzi, R., Dos Santos, T. B., da Conceição, D. M., Rodrigues do Nascimento Jr, D., ... & Coiffard, C. (2023). A new remarkable Early Cretaceous nelumbonaceous fossil bridges the gap between herbaceous aquatic and woody protealeans. Scientific Reports, 13(1), 8978.
Li, Y., Smith, T., Svetlana, P., Yang, J., Jin, J. H., & Li, C. S. (2014). Paleobiogeography of the lotus plant (Nelumbonaceae: Nelumbo) and its bearing on the paleoclimatic changes. Palaeogeography, Palaeoclimatology, Palaeoecology, 399, 284-293.
Matthews, P. G., & Seymour, R. S. (2006). Anatomy of the gas canal system of Nelumbo nucifera. Aquatic Botany, 85(2), 147-154.
Vogel, S. (2004). Contributions to the functional anatomy and biology of Nelumbo nucifera (Nelumbonaceae) I. Pathways of air circulation. Plant Systematics and Evolution, 249, 9-25.
巽二郎, 野田, 千代, 山内, 章,飯嶋, 盛雄, & 矢野勝也. (1999). ハス (Nelumbo nucifera) 葉身における表面温度の分布特性. 日本作物学会東海支部会報, 15-18.
Blaylock, A. J., & Seymour, R. S. (2000). Diaphragmatic nets prevent water invasion of gas canals in Nelumbo nucifera. Aquatic Botany, 67(1), 53-59.
https://www.tsukuba.ac.jp/community/students-kagakunome/shyo-list/pdf/2018/j1.pdf
水に潜ったマコモ Zizania texana
みなさんお楽しみの「世界の珍妙水草」シリーズです。今回は、身近な植物に近縁ながらもどうしてこうなった??という、イネ科を紹介しようと思います。
日本人にとって馴染み深いイネ、およびイネ属Oryzaは、熱帯の雨季と乾期に劇的に水位が変わる環境に特化した、まごう事なき水草といえます。イネを栽培していて、よく思うことがあります。「イネは沈水できるのか?」と。
結論から言えば、現状では否と考えています。イネを深い水深に蒔くと、CO2添加が多くとも少なくとも結局水面に到達できなければ枯れてしまう…というのが私の試した限りでの感想です。浮稲や一部の野生イネのように水面を突破するまで激烈な成長が見られるものや、逆に一部の野生イネや一部の洪水耐性稲のように成長をピタリと止めて耐え凌ぐものはあれど、沈水状態で旺盛に生育するイネ属は今の所、私の耳には入ってきていません。
では、イネ属の近縁属ではどうでしょうか?
その作物種が無理でも、近縁な他属の遺伝資源を利用することはよく行われ、例えば小麦では根の水没に耐えるミズタカモジの遺伝資源が注目されて、ハイブリッドが作られたりしています。(実用化には至っていませんが…)
将来的に工業的な栽培を行うにあたり、「沈水できるイネ」いや、沈水で旺盛な生育を示すイネ」は大きな需要があるはずだと確信しています、しますよね(強制)
↑私が見たいだけである。
イネ属Oryzaに近いものといえば、かつてイネ属に含まれていた数属やヒグロリザHygroryza、マコモ属Zizania、あとはサヤヌカグサ属Leersiaあたりでしょうか。そのなかに沈水性のものがあれば、もし物好きな人がいれば細胞融合等によって沈水イネを作出でき、将来的に水中で工場的にイネを培養できるかもしれません。ヒグロリザは浮くイメージが強いですが、全ての水上部が刈り払われると一時的に薄く繊細な沈水葉をだして生育します。これはイネ科としては珍しい明瞭な水中異形葉ですが、旺盛に生育する、というのとはまだずれているでしょう。サヤヌカグサ属はまた後にするとして、マコモ属はどうでしょうか?
マコモといえば日本人にもお馴染みの植物で、種子は菰米として日本においても中国においても古代から利用されてきた植物です。その後マコモダケが利用されるようになり、今ではそちらの方が有名ですが…。北米では近縁種のZizania aquaticaとZizania palustrisがネイティブアメリカンにより古くから採集利用され、こちらは現在もワイルドライスとして利用されています。
さて、これらマコモ類は抽水性植物として水辺に茂っているイメージがありますし、野外でも抽水性で見かけることがほとんどです。しかし日本のマコモにおいてすら、なんというか沈水ポテンシャルがあるのではないか?という気がしないでもありません。マコモは下流域の泥底に出てくるイメージが強いですが、見ていると湧水の影響をかなり強く受ける場所にも分布のピークがあり、しばしば最優占種となります。そうしたところでは、湧水による何らかの恩恵を受けてマコモが優占しているのではないか?とすれば実生の沈水生育では?と、思ったこともあります。しかしながらマコモの実生栽培は種子が乾燥に弱いため保存が難しく、収穫適期が短く、種子の休眠が強いことからなかなか難しく、当方では思うように実証実験をできていません。さらに実生のマコモを野外で識別することもなかなか難しいことから、野外観察でも謎のままでした。
ワイルドライスは一年生で、しかもマコモ並みかそれ以上に深い水深から生えてくることから少なくとも止水におけるかなりの水没耐性は持っていそうです。しかしながら、私の調べた範囲ではワイルドライスにしても、マコモにしても、水中での良好な生育を示したという報告はなく、あくまで抽水性植物としての扱いでした。
しかし北米には、なんと開花結実以外を水中で完結させるマコモが存在します。
それが今回紹介する、Texas Wild Rice テキサスワイルドライス Zizania texanaです。
テキサスワイルドライスは英語圏ではそこそこ知名度のある植物らしく、英版Wikipediaが大変充実しています。しかし日本語の文献でこの珍妙な水草に触れたものは見たことがないので、耳慣れない方がほとんどでしょう。
この種を初めて見つけたのはテキサスの弁護士でアマチュア植物学者でもあったW. A. Silveusであり、1932年に見つけた当初はノーザンワイルドライスZizania palustrisと考えたようです。しかし記述にあるものとは異なり川幅40フィートの急流の流れる川底に1~4フィート沈水して生育していました。水位が急に上昇して草が水没したとすれば理解できる状況でしたが水位は常に高い環境であり、流れの周囲にも抽水性の個体は見られませんでした。また彼は葉は長さ5フィート、幅は基部で8~10㎜、上部で15~20㎜、花の付いた部分は1~2フィート水面から突出して生育し、水のない部分では茎は鋭く屈曲してときに節から発根していたと記しています。その後花期が4月から11月にもおよぶことを発見しています。これらの内容を記した手紙と草体の生態写真はA. S. Hitchcockに送られ、1933年にHitchcockにより記載されました。
本種はエドワーズ帯水層に灌流される、サンマルコス川の上流域に生育します。湧水の影響で水温は年中20度前後(8月で25.5度、2月で20.4度)、水源が石灰岩を通過することからpHは7.4~7.9と弱アルカリ性のようです。沈水状態で発芽生育し、幅15mmほどの葉は長さ1m、茎の長さは数メートルまで成長し、切藻増殖します。日本のマコモに慣れていると意外ですが、マコモ以外のマコモ属はこの種以外は全て一年草であり、切藻増殖して多年化するのは湧水への適応により獲得した形質であるのではとも連想されます。かなり深いところにも生育できるようで普段は水中に生育し、水面まで到達することすらあまりないようです。そのため、水位が減少したタイミングで水面に花をあげて開花します。このような湧水依存の生態を身につけたのは豊富な湧水量によるものと思われますが、地下水の灌漑により湧水量が減少し、発見当初こそ大繁茂して灌漑の障害となったもののその後激減し、1970年にはほぼみられなくなりました。1976年に2.4㎞の範囲で再発見されたものの、いまも3km程の範囲にしか現存が確認されていません。
個人的には、1978年の調査でPotamogeton ilinoensisやVallisneria americana、Sagittaria platyphyllaが同所的にみられたのは当時現存していた2.4㎞のうち上流端の0.4㎞であり、下流側の2㎞ではテキサスワイルドライスが優占種となっていたことはきわめて興味深いです。というのも、イネ科植物が湧水で優占するのは通例は湧水の最上部であり、トチカガミ科やヒルムシロ科の沈水植物は一般的にその下流側で発生するのが通例のように思われるためです。イネ科においてここまで湧水に依存し、生育期間のほとんどを沈水状態で過ごす種はほかに、パッと頭には浮かびません。
本種はいまのところ生息域においてはかなり大きな現存量がありますが、その生息域は極限され、地下水利用が止まらない限り湧水量は減り続けることでしょう。
この変な水草が残り続けてくれることを願います。いつか農業の役に立ったり…しないかなぁ。
ヒルムシロ属のSclerenchymatous rays
この形質はエゾヤナギモ種群、およびヤナギモ、P. polygonus、P. ochreatusにみられるとされるが、北半球でみられるのはエゾヤナギモ種群とヤナギモのみ。

